
So the other day I started the task of downloading all the files marked “Ornithoscelida” that are currently my brain and putting them into some sort of logical order (sort of). My first post on this topic just quickly outlined some of the reasons why there is some controversy now looming within my field in terms of our understanding of the origin and early evolution of the Dinosauria… (which is the title of my PhD thesis, by the way).
Whatever the outcome of the Langer et al. 2018 reply and our rebuttal in terms of the data matrices and trees produced, I have a strong hunch that the traditional model of early dinosaur evolution is not accurate.
And.
Here.
Is.
Why… (queue wild speculation!)
Firstly!
Ornithischia – Saurischia as a dichotomy fundamentally dividing all dinosaurs into two distinct and neat piles seems to me to be a little… shall we say, overly simple.
The idea was based upon some observations made by the British Palaeontologist Harry Govier Seeley back in the 1880s. Now, the crux of his argument was centred on the construction of the hip bones in the two ‘groups’, and the pneumatic features (or lack thereof) within the bones of the axial skeletons.
The hip thing was focused upon when Seeley picked out his names: Ornithischia = ‘bird-like hip’; Saurischia = ‘lizard-like hip’.
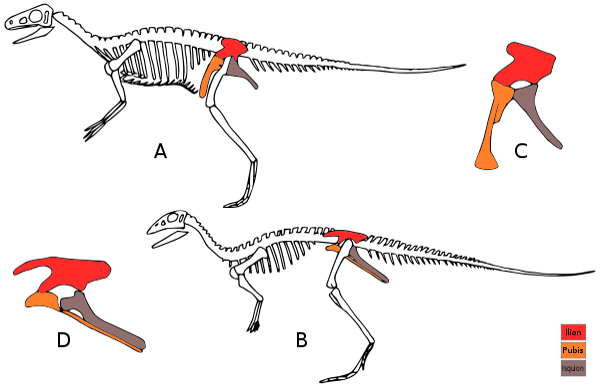
As implied, the hips of saurischians is much more like that of most other lizards and raptiles, with the pubic bones (pubes, yes, you read that right) facing forward, into the space between the hind limbs and abdominal cavity.. Ornithischians, on the other hand, have retroverted (pointing the other way) pubes, creating a space between the hind legs, and this condition is also what we see in birds, hence the name.
Now, while that might seem significant, it is worth noting that within the group of animals that we today, and Seeley in 1887, would call the dinosaurs, there are actually three distinct groups that we can fairly assume are monophyletic (fancy science word for all descend from the same common ancestor) – Ornithischia, Sauropodomorpha and Theropoda (here arranged in terms of how much worth I give each of them).
Saurischia, as termed by Seeley, grouped together the Theropoda and Sauropodomorpha clades into one larger clade, based upon the fact that they appeared to share the same hip structure and skeletal pneumaticity.
However, given that we know that the hip structure is in fact a “primitive condition”, meaning it is a feature that they share simply because they haven’t changed from the ancestral state, we actually cannot use it as a feature to group Theropoda and Sauropodomorpha… It’s primitive, and therefore not derived, and we only group organisms based upon shared derived features (synapomorphies, in the technical jargon).
Why?
Because just because a subgroup have gone on to lose a feature that was the ancestral condition for the wider group, it does not mean that we can then say that the other subgroups who have ‘hung on’ to the feature should be grouped together to the exclusion of the experimental group, at least based on that feature’s absence/presence, without other evidence.
An example.
Cetaceans, for those of you who are unaware, are the group of mammals that includes all living whales, porpoises and dolphins, as well as many of their extinct close relatives. Now, if we were to look at three mammalian groups, say, Cetacea, Carnivora (bears, dogs, cats, hyenas etc.) and Primates (us and monkeys and stuff), and tried to group them based upon a single choice feature, lets say ‘feet’, we would get an incorrect tree.
Whales and dolphins obviously lack feet. Primates and Carnivores have feet.
Does this mean we group Primates and Carnivores together to the exclusion of whales and dolphins? Well, no, is the answer.
We know, from a pile of other anatomical (and, I imagine, also genetic) evidence that the possession of feet is a feature in mammals that they all inherit from their common ancestor (and from the common ancestor of all tetrapods to be fair). Now, just because the group that contains whales and dolphins decided, for whatever reason (my hunch is that it was their aquatic lifestyle), to abandon feet, this doesn’t mean that they should be the ‘outgroup’ to a monophyletic clade containing only carnivores and primates.
In fact, Cetacea is more closely related to Carnivora than either group are to the Primates. Like I said before, you need to look at TOTAL EVIDENCE to come to this quite obvious conclusion, which means focusing on more anatomical evidence. But, more importantly, the features you pick should not be those features which, from the fossil record and phylogenetic bracketing, we know are only in certain subgroups because everything within the larger group would have it as a ‘default’ if it wasn’t for later, specialisations in some subgroups.

Anyway, that’s all well and good for the hip problem, but “WHAT ABOUT PNEUMATIC FEATURES MATT!” i hear you scream while tightly gripping your little collection of ‘saurischian’ plush toys, including your fluffy Velociraptor and your very-much-not-life-sized Brachiosaurus.
Well, I’ll get to that too… now, I guess…
So, skeletal pneumaticity is, essentially, the reduction of parts of the skeleton, usually the axial (spinal) column, to reduce weight (maybe) and to fill with air sacs. And, understandably, it was the other major feature that Seeley pointed to as being unique to saurischian dinosaurs (i.e. not seen in ornithischians). If you would like to read more about this feature, head this way.
Anyway, it is certainly true that many derived theropods and many derived sauropodomorphs (actually, its mainly ‘true sauropods‘) have such pneumatised skeletons. The question is, is this feature a shared derived trait, taken from the common ancestor of these two groups that did not also give rise to the ornithischian lineage? Or, did this feature appear convergently within the two subgroups?
As often happens in the history of evolution, the same kind of features appear in different, unrelated groups. And by unrelated, I mean far apart enough on the tree that we can be confident that the feature that they possess has appeared separately, by coincidence, for a similar function, rather than from an ancestral state. Birds’ and Bats’ wings is a classic example of two unrelated (very distantly related) groups evolving functionally similar features.
Could this be the case with the pneumatised skeletons of the big tree browsing sauropods and the large meat eating theropod dinosaurs?
It is true that for Seeley, the material available to him was limited when compared to what we, as modern palaeontologists, have to work with. The rates of new dinosaur discoveries have sky rocketed in the last few decades and we now know far more about all the major groups of dinosaurs than we did even when my PhD supervisors were students. Especially from within the Triassic Period, the first of three periods in which we see non-avian dinosaurs, we have increased our knowledge in a very dramatic way since the late 1800s.
Seeley was right when he observed that many of the Late Jurassic and Cretaceous sauropods and theropods both had ‘hollowed out’ bones. What he did not, and could not, know at the time was that perhaps, way back toward the origins of these two groups, the features would become less well developed and in some, very early specimens, be absent entirely.
If you go way back, as in way, way back, to the roots of Sauropodomorpha and Theropoda, what we see are distinctly less pneumatised skeletons in the ‘basal’ forms, with animals like Pantydraco, a basal sauropodomorph, completely lacking any kind of pneumatic features. Could this be a reversal in this taxon, which is a derived feature and a departure from the ancestral ‘saurischian’ condition of having some pneumaticity? Or is this an indication that those features we see, and Harry Seeley saw, in much later appearing, highly derived and specialised sauropodomorphs and theropods, not common but convergent? Skeletal pneumaticity seems to reduce and then dry up completely as we trace the lineages back from the big old beasts of the Jurassic and Cretaceous to the Late Triassic critter that have been discovered since Seeley’s great divide was erected.
For me, looking at lots of the earliest examples of ‘saurischian’ dinosaurs, and noting the progression of the pneumatisation in the diverging lineages, I am convinced that the massive holes we see in the axial columns of sauropodomorphs and theropods is an example of convergence.
I think that Seeley’s observations were good, for their time, but perhaps need to be reassessed in the light of a huge amount of fresh new evidence, gleaned from the fossil record more recently.
Then there is yet another feature that adds complexity – pterosaur pneumaticity, and pneumatic features in animals outside of Dinosauria.
Recent discoveries of close dinosaur relatives, which fall on the line within Archosauria that leads to birds and not to crocs, seem to suggest that there might be more to this problem than we though. Some of these new creatures seem to have some features indicative of partial skeletal pneumaticity, or, at least, some features that are ‘laying the groundwork’ for more complex pneumatic recesses and spaces in the skeleton in later, dinosaurian forms.
Now, if these features are homologous with what we see in Saurischia, then we run into exactly the same problem as with the hips.
We have already seen how Shakira is perhaps wrong on this one point – that hips do lie – but perhaps this same logic extends to pneumaticity. If ornithischians lack it, but it is actually the ancestral condition that would have been the ‘default’ for all dinosaur subgroups, then the ornithischians’ awkward choice to reject the feature doesn’t tell us squat about where they fit in the tree… its dolphin feet all over again.
There are hints of possible pneumatic features in some ornithischians, such as heterodontosaurus, pictured below. In its skull there are, as has been observed by some, recessed similar in nature to the pneumatic features of certain theropods and basal sauropodomorphs.
Whatever the case, I think for now the pneumaticity evidence for a monophyletic Saurischia should be treated with extreme cation.
When we scored as many independent characters as we felt we could, for as many species as we could, including a whole host of Triassic and Early Jurassic species that were simply unknown to the world during the age of the likes of Seeley, we found that the total evidence, when analysed, rejected the age old concept that the ornithischians must be the ‘odd outgroup’ to a monophyletic Saurischia.
Langer et al.’s response showed that the alternative arrangement, that preserved the traditional model, was not statistically significantly different to our own hypothesis, and that was with much of our data having been altered, in ways that we perhaps disagree with strongly.
Interestingly, some of the characters that were very frequently re-scored by the other team, were those that pertained to pneumatic features.
As I said in the previous post, much more needs to be done to address these current areas of disagreement.
In the next post, I will endeavour to lay out more about my alternate view of Ornithischia, with the evidence that I have been observing myself since our original publication.
For now, let me just close by saying this.
If, by some miracle, by some Divine insight, or bizarre circumstance that is beyond all human reason, a creationist (yes, Seeley did not even believe that evolution was a thing), a creationist with relatively little to go off (compared to today’s researchers), who only looked at late and derived forms, and didn’t apply the principle that synapomorphies trump plesiomorphies, made, in 1887, an observation that is still scientifically valid, I will eat my shoes!
If, somehow, that fundamental dichotomy, which runs from the very tips of every dinosaur lineage, dead and alive, all the way down to Dinosaur 1.0, the most recent common ancestor of all dinosaurs, I’ll melt down my belts and make a sauce to pour over my shoes, which I am eating.
It would be absurd to think that, sometime in the Middle (or possibly Early) Triassic, two descendants of the first true dinosaur separated and, one following mum and keeping its hips just like all its other reptile friends, and one (the black sheep of the family) retroverting its pubis in an attempt to break free of the mould that had held back so many promising young dinosauriforms before it, gave rise to the two groups that Seeley, A CREATIONIST with EXTREMELY LIMITED DATA has supposed, mused, guessed at, from the comfort of armchair in King College, London over 130 years ago… It would be crazy if he was right.
Even more remarkable still when we consider that the Triassic fossil record for one of these two ‘fundamental groups’, the Ornithischia, is basically non existent.
Maybe he was right, or maybe we’ve had it wrong and Seeley’s work was just a best guess with what he had at the time, but either way, I think that it is certainly not something that we should just take as a given, and I think that a lot more work needs to be done if we are ever going to truly solve the mystery that is the early evolution of the dinosaurs.
I for one am excited about what the future holds in this particular area of palaeontological research.
by
Dr Matthew Grant Baron

2 thoughts on “Thoughts on Ornithoscelida … over one year on … (part 2)”
Comments are closed.